字體:小 中 大
字體:小 中 大 |
|
|
|
| 2026/07/06 21:35:27瀏覽358|回應0|推薦1 | |
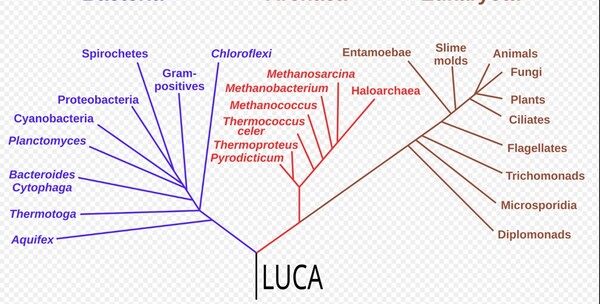
我們與地球上的所有的幾百萬種生物是從哪裡來的?近代演化理論說是『它』! 慎終追遠:追、追、追……不只追到『南方古猿(Australopithecus)』,哺乳類祖先『合弓綱(Synapsids)』!這些演化學者都覺得還不夠遠。談到地球上所有生物起源的『最後共同祖先LUCA』,照近代廣進化理論:這個42億年前的祖先LUCA最終演化成地球上的幾百萬個物種……哇嗚!這可不是一件小事!能不多了解它一點嗎? 『最後共同祖先LUCA』我們大家都從它演化來的? 談到近代演化理論所主張——親緣關係樹基部的最底層,亦即演化理論中太古時代在真細菌域與古菌域之前的最後共同祖先—(LUCA;Last Universal Common Ancestor),我們要先談到達爾文在 1859 年出版的《物種起源》(On the Origin of Species)中,於第十五章寫下了著名的論點,這似乎表達了達爾文本人關於LUCA的類似觀點(註1): 「依據伴隨着性狀分歧的自然選擇原理,動物和植物從這些低等的中間類型發展出來,並不是不可信的;而且,如果我們承認了這一點,我們必須同樣地承認曾經在這地球上生活過的一切生物都是從某一原始類型傳下來的。」 換句話說,LUCA演化生物學推導出來的假設,指地球生物最原始的共同祖先,是地球上所有現存生命的共同起源,但這共通祖先未必是最早的生命。進化論者一般相信,最後共同祖先在古太古代出現,距今約42億年。最後共同祖先分化出細菌與古菌,演化成各種生命(註2)。在演化生物學中,因為科學家無法無法挖到35億年之前的細胞化石,因此若要拼湊出演化理論上分子生物學的『最後共同祖先—LUCA』的可能面貌,完全仰賴一套一廣演化概念設計精密的生化計量學與大數據技術,如:使用核糖體核糖核酸(rRNA)來予以推斷(註1)。演化論學者所提及的LUCA被認為是一種「標準化、較為現代化」的單細胞級生物物種,已經擁有完美的雙股 DNA、形成通用的64個RNA密碼子,以及與現代生物一模一樣的核糖體工廠,具有精密的F-ATP分子馬達,並擁有了跨越細胞膜運輸的通道蛋白系統。這就是使用「分子回溯或親緣關係重建(Molecular Retrodiction / Phylogenetic Reconstruction)」而試圖勾畫出最後共同祖先的「樣貌」(註8)。但以上所列出的這些『生命的要件』卻顯示這樣貌沒有一項是簡單的,而且全都是『極複雜的分子機器』!
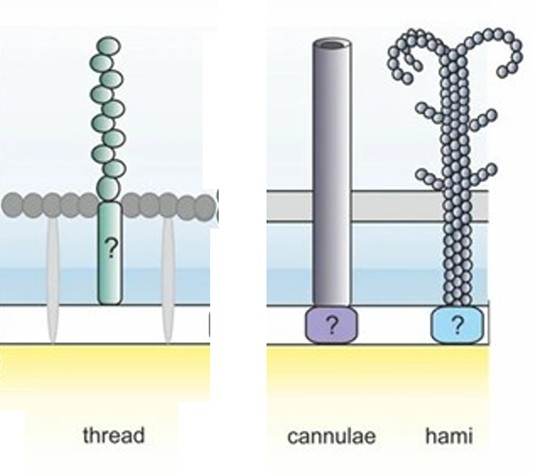
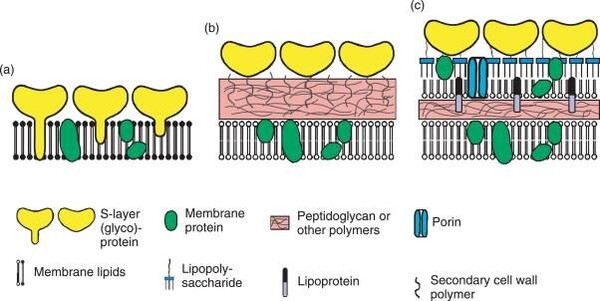
圖2-7 1990年的系統發生樹,其中將生物所有的主要類別都連結到了最後共同祖先(LUCA),而這張圖的分類依據是核糖體核糖核酸序列。圖片來源:維基百科關鍵字『最後共同祖先』,由Chiswick Chap-自己的作品, CC BY-SA 4.0 而"LUCA" 這個現代人朗朗上口的縮寫,是在1994年被正式創造出來的。提出者是法國著名的進化微生物學家帕特里克·福泰爾(Patrick Forterre)。在1994年於法國舉辦的一場關於「早期生命演化」的國際學術研討會上,福泰爾為了方便與會科學家溝通,正式提出了LUCA這個縮寫。這個名字不僅好記,而且在法語和許多歐洲語言中,"Luca" 也是一個常見的男子名(源自拉丁語 Lucas,意為「帶來光明的人」),這個擬人化的雙關語讓這個科學概念迅速在國際學界與大眾科普中流行開來。LUCA之前仍被演化論者認為可能有更古老的共同祖先,但是這場以演化論觀點跨越古生物共同祖先LUCA的時空鑑識工作,主要是透過以下四個層面的步驟與方法進行的(註3;註4): 1. 篩選「全域同源基因」(Universal Homologs):電腦會自動剔除那些只存在於少數物種的特異基因,專門篩選出同時存在於真細菌域與古菌域兩大領域中的「核心基因」。經過極其嚴格的篩選,科學家最終鎖定了大約 355 個核心基因群(Clusters of Orthologous Groups, COGs),這被認為是 LUCA 必定擁有的「最小基因集」。 2. 多重序列比對(Multiple Sequence Alignment, MSA)與概估的分子鐘(Molecular Clock)「起點時間」——進化論者進而推算出樹根主幹 LUCA 活在距今大約 41 至 43 億年前 的冥古宙。 3. 三維結構同源性預測(3D Structure Alignment):科學家不再只比對「DNA 字母」,而是比對真細菌域與古菌域兩大領域中的蛋白質折疊出來的「3D 空間形狀」。如果細菌的某個代謝酶與古菌的某個酶,其 3D 折疊結構(Fold Domain)幾近完美重合,分子生物學家就能突破序列的限制,斷定這個結構域在 LUCA 時代就已經成型。 4. 經由生態學與生化路徑「反推」生活環境:分子生物學家針對LUCA的355 個核心基因群開始進行「功能分類(Functional Annotation)」,看這些基因到底是負責做什麼的:分析認為第一類是依賴鐵、硫、鎳、鋅等金屬簇(Fe-S clusters)的金屬酶基因。第二類是氫氣與二氧化碳的 Wood-Ljungdahl 代謝途徑基因。第三類是耐熱酶(如 Reverse Gyrase,反向旋轉酶,通常存在於嗜熱菌中)。 正如2024慕迪所率領的研究團隊所述(註4): 『系統發育分析表明,LUCA的基因組大小至少為250萬鹼基(249萬鹼基至299萬鹼基),編碼約2600種蛋白質,與現代原核生物相當。我們的研究結果表明,LUCA是一種原核生物級別的厭氧產乙酸菌,並擁有早期免疫系統。』 儘管LUCA經演化生物學家是如何透過程式碼、生化計量與分子回溯,在電腦中模擬出 42億年前 LUCA 的生動樣貌,說起來似乎言之鑿鑿,但LUCA的『假定存在』卻暴露出演化理論本身的重大矛盾。這些重大矛盾很少被演化論者談到! LUCA的細胞膜是哪一種?是哪種都不對!LUCA—最後共同祖先的矛盾 LUCA—這個演化學者口中的最後共同祖先其理論背後最重要的核心假定,以及它們在科學上面臨的質疑有: 1.真細菌域的細胞膜的基本化學構成是『雙層磷脂質』;古細菌域的細胞膜則是『單層/雙層甘油醚』(如圖2-3)。共同祖先LUCA的細胞膜是哪一種呢?變成是哪一種都不對! 演化學的學術論文對此進行了鋪天蓋地的瘋狂討論,且至今都沒有絕對的定論。這就是演化生物學中著名的「脂質矛盾(Lipid Divide)」!如果』我們去比對現代生物的細胞膜,會發現一個根本性的化學對立:細菌與真核生物: 細胞膜是雙層磷脂質(甘油-3-磷酸 G3P的酯鍵脂肪酸長鏈)。古菌(Archaea): 細胞膜通常是單層/雙層甘油醚、G-1-P手性、具有支鏈異戊二烯鏈,有些高溫古菌還會融合成跨膜單層四醚結構)。既然兩者的細胞膜是手性相反(鏡像對立)、代表製造細胞膜的合成酶完全是不同源的結構,演化論學者的學術論文針對「LUCA 的細胞膜長怎樣」主要分裂成了四大假說派別。其中最主要的派別Caforio, A., et al.團隊 (2018)主張LUCA同時含有酯鍵和醚鍵、G3P與G1P混雜的「雜交混合膜」細胞。理論根據是:在實驗使用基因轉殖的方式,把古菌合成甘油醚膜的酶基因,植入大腸桿菌(細菌)中,可以成功製造出了同時含有酯鍵和醚鍵、G3P與G1P混雜的「雜交混合膜」細胞(註5)。 以上演化論學者Caforio, A.團隊的實驗假定其實是使用了『基因轉殖』此種最『非自然』的人工方式來『強迫』兩種不同結構的膜『硬是要』合而為一。生物科技公司基因轉殖人類的胰島素基因也可以使大腸桿菌製造出多種人類的胰島素,但人類與大腸桿菌兩者卻是大相逕庭的。因此能夠把古菌的基因轉殖給大腸桿菌這又代表甚麼呢?能證明這兩類生物很相似嗎? 2.再者演化論學者也刻意跳過了最棘手的真細菌與古菌細胞膜鑲嵌的『膜蛋白系統如何能彼此吻合?』的部分。上過高中生物課的人都知道細胞膜的基本構成是流體鑲嵌模型(Fluid Mosaic Model),也就是細胞膜上鑲嵌了四大類蛋白質:1.管制各式分子進出的通道蛋白(形成細胞膜有半透性);2.代謝的酵素;3.辨識作用的醣蛋白與寡糖脂;4.對外進行化學訊號溝通的受體蛋白(用以感測外界環境變化,如前方有食物)。不同類的古菌與真細菌豈是只有把化學上不同性質的膜混合為一的問題而已?古菌與真細菌兩類生物細胞膜上『整套蛋白質機器的機件(鑲嵌的膜蛋白)』都是徹底的不同!根本就是如同兩種不同性質的工廠:自小客車零件工廠(真細菌)與越野車零件工廠(古菌),兩者本質上差很多且無法相容! 如果LUCA的細胞膜是A(真細菌—雙層磷脂質)與B(古菌—單層甘油醚)混雜型的機件形成的機器,其機器是要如何能運轉呢?賓士汽車與福特汽車兩種不同車系的零件可以相互組成『拼裝車』嗎?而他們難道沒有可能就是根本沒有演化關係的兩個起源,根本不需刻意以『均是出於LUCA共同祖先』這樣演化概念予以刻意連結而將LUCA硬塞入進化模式『一棵親緣關係樹』的框架之內嗎? 在凱西.盧斯金的專文也提到進化論學者庫寧(Eugene V. Koonin) 對『細胞起源』與『細菌及古菌起源』分子生物學分析的一段絕佳評論(註6),印證了演化論者心目中的最後共同祖先LUCA的「樣貌」,其實充滿分子生物學的不確定性: 『細胞的起源 古菌和細菌是兩種主要的細胞類型(也是生命的兩大原核生物域),它們具有化學成分不同的細胞膜、主要不同的膜生物合成酶以及不同的核心DNA複製酶。這大大增加了重建古菌和細菌細胞祖先的難度,並提示我們需要尋找其他解決方案。 細菌和古菌主要分支(門)的起源 儘管細菌和古菌在同一域內的分子一致性遠高於域間一致性(特別是,膜和複製機制在各域內都具有同源性),但古菌與特別是細菌系統發育樹深層分支分子空間結構的拓撲結構仍然難以捉摸。這些親緣關係樹在分析的基因和所採用的方法方面明顯缺乏穩健性,而且儘管人們已付出相當大的努力來界定細菌的高級分類單元,但達成共識的跡象仍然不明顯。古菌分為兩個分支-廣古菌和克倫古菌-的劃分較為明確,但即使是這種劃分也未必能在親緣關係樹中得到體現,古菌域的進一步劃分仍然模糊不清。』 3.雖然都是有附著功能的菌毛結構,古菌菌體表面的彼此互相連結的線狀菌毛(thread)、鉤狀突起(Humi)或套管(Cannulae)與細菌菌體表面的線毛(Pili)在生化結構上兩者截然不同(註7;註8;圖2-8)。細胞膜外側的細胞壁(古菌—S層【S layer】或假肽聚糖;真細菌—肽聚糖;圖2-9) 兩者分子生物學的相似性也是天差地遠。這造成LUCA作為真細菌域與古菌域親緣關係樹『單一演化起源』基幹的極端矛盾。
圖2-8 古菌菌體表面的彼此互相連結的線狀菌毛(thread)、鉤狀突起(Humi)或套管(Cannulae)與細菌菌體表面的線毛(Pili;圖左)在生化結構上兩者截然不同(註13;註14)。細胞膜外側的細胞壁(古菌—S層【S layer】或假肽聚糖;真細菌—肽聚糖) 兩者分子生物學的相似性也是天差地遠。這造成LUCA作為真細菌域與古菌域親緣關係樹『單一演化起源』基幹的極端矛盾。圖片來源:(左)https://www.sciencedirect.com/science/article/pii/S000634959877821X (中、右)https://link.springer.com/article/10.1007/s40588-023-00204-7
圖2-9 圖(a):某些古菌具有甘油醚細胞膜與S層蛋白質外覆結構的細胞壁 圖(b)革蘭氏陽性菌細胞膜:具有厚的肽聚醣外層細胞壁 圖(c)革蘭氏陽性菌細胞膜:具有薄的肽聚醣外層與甘油磷脂雙層的外膜的複合細胞壁 圖片來源:https://boku.ac.at/en/btlw/nabi/research/generation-and-characterization-of-artificial-archaeal-cell-envelope-structures-and-their-relevance-as-model-membrane-platforms 從演化起源(Evolutionary origins)來看,古菌的鉤狀突起和真細菌線毛分子結構甚至並非同源構造,組裝機制的分子生物學差異極大!許多古菌的表面延伸物(包括古菌鞭毛 Archaellum 以及部分古菌線毛)雖然在組裝模型上,普遍被認為與細菌的 第IV 型線毛(Type IV Pili) 類似(由基部向外推擠組裝),但古菌體表的鉤狀突起的主要組成蛋白(約 120 kDa 的次單元)與細菌線毛蛋白(Pilins)的基因同源性極低。2005年的研究(註6)指出確認古菌體表鉤狀突起的刺網狀主幹(直徑約 7–8 奈米),且其末端具備 60 nm 的三叉倒鉤結構,能在極端環境下維持高強度的化學與物理穩定性(耐受 pH 0.5–11.5,溫度 0–70°C)。古菌的鉤狀突起最初是在一種生活於冰冷、富含硫化物泉水中的特殊古菌(如與細菌共生的 SM1 euryarchaeon)表面被發現。這種極其複雜的「倒鉤」構造(見圖2-8),演化論學者認為是古菌為了在特定的流體動力學與極端生態棲位中,確保自身與共生夥伴不被沖走、維持群落結構而演化出的獨特適應策略。但是奇怪的是:細菌的線毛與古菌的鉤狀突起屬於分子生物學『結構不同卻功能相同』的趨同演化(Convergent evolution)的典型例子,彼此屬於與演化原理牴觸的『同功構造』(見圖2-8)!它們在「協助細胞附著與建立群落」的功能上類似,但古菌體表鉤狀突起在末端演化出了不可思議的物理倒鉤與主幹刺狀結構,在機械精密程度上,相對單純的細菌線毛與古菌不可同日而語!但是奇怪的是:真細菌與古菌這兩類的生物卻被進化論者認為有共同祖先LUCA!硬是接上相同的演化起源以求符合演化的『單一起源假設』!像這樣削足適履,硬把兩個截然不同結構的『主幹』硬是套在『同一棵』親緣關係樹上,請問這樣的廣進化邏輯是合理的嗎? 對以上假說,智慧設計論學者結構生物學家安·高哲(Ann Gauger,麻省理工學院學士、哈佛大學博士後研究員)在她的研究與評論中也指出重要的關鍵點(註9): 『細菌的 G3P酯鍵和古菌的 G1P 醚鍵是絕對的呈現“立體化學鏡像”對立‧……酵素(筆者註:生物的各種代謝需要藉著不同類別的酵素)作用的活性位點(Active site)對受質(substrate)的立體手性要求是極致嚴苛的。在工程學上,你不可能透過「隨機改變(如同基因突變)一個零件」,讓一個組裝左手螺絲的工廠變成組裝右手螺絲的工廠,其中間型態只會製造出廢料!因此,「混合膜」在現實生化反應中是不可能順利與現代膜蛋白協同演化的。』 4.嚴重邏輯錯亂:演化學者承認最後共同祖先LUCA是一個42億年前遠古,卻擁有2,600個基因的極複雜結構!LUKA既然被推論在演化架構下是42億年前產生,卻必須要有至少2,600個基因來編碼蛋白質?試想:LUCA這樣遠古的演化祖先可能『極古老又極精細』嗎? 即使用膝蓋加以想像,都不難發現這與達爾文『由簡趨繁』的的演化觀點是有嚴重衝突的!正如凱西.盧斯金所言(註10): 『這就引出了一個問題:生物體生命,即“非常複雜的細胞”,是否可能在低於該研究歸因於 LUCA 的複雜性閾值(筆者註:低於生命可存活許可複雜度的最低水平)的情況下存在——例如,如果沒有像“對抗病毒的免疫系統”或“抵禦紫外線損傷的基因”這樣的複雜防禦機制。 結構簡單的細胞是否能夠存活?這種想法本身就有點匪夷所思。如果結構簡單的細胞能夠存活,那麼就需要有人解釋這種毫無防禦能力的生命體如何能夠生存和繁殖。如果結構簡單的細胞無法存活,那麼就強烈暗示生命可能起源於非生命物質,而這原始生命卻完全沒有經過一個結構簡單但同樣能夠存活的過渡狀態,這樣的邏輯顯然是矛盾的。(筆者註:生命怎麼可能從「非生命」直接跳躍到 「高度複雜且能存活」的細胞?這機率低到像神蹟)』 論LUCA若真實存在的兩個基本前提 而進化論者自己也承認LUCA若是真的存在需有諸多基本假定都要成立,例如: 前提1:真細菌域與古細菌域的微生物分子回溯中的「垂直遺傳(VGT;Vertical Gene Transfer)」訊號,必須沒有被它們的「水平基因轉移(HGT;Horizontal Gene Transfer)」完全抹去。演化論學者所主張藉分子生物學的DNA基因序列分析,可以透過比對細菌和古菌共有的 355 個基因簇 (約2,600個基因編碼蛋白質)來重構 LUCA。這背後有一個巨大的數學假定:這些基因在過去30多億年間,主要的是透過「代代相傳(垂直遺傳)」留下來的,科學家可以用統計學把它們過濾、辨識出來。 細菌的垂直遺傳指的就是透過分裂生殖的過程(主要是二分裂法,Binary fission即一隻分裂為兩隻),將基因傳給下一代的過程。如果用人類的社會來比喻:垂直遺傳(如果把DNA的傳遞形容成財產的移轉),垂直遺傳就像「傳家寶」,是父母(親代細菌)把資產(整套基因DNA訊息)代代相傳給自己的子女(子代細菌)。水平遺傳(接合作用、轉形作用等):就像「鄰居借(送)東西」,是同輩之間(甚至是不同種細菌)互相交換(買賣)隨身碟(質體)。 1. 尋找目標與鎖定(Recognition & Attachment):擁有特定也就是說,水平基因移轉就像細菌有線毛(Pili,特別是其中的性線毛 Sex pili)的結構,線毛之所以能夠交換遺傳物質,靠的並不是像管子一樣把 DNA「吸」過去,而是扮演了類似「拋繩槍」與「接駁橋樑」的角色。這個過程在生物學上稱為接合作用(Conjugation),是細菌進行基因水平轉移的重要機制。以下是它運作的詳細步驟:DNA質體(例如 F+質體,含有製造線毛與轉移基因的DNA訊息)的供體細菌(Donor),會向外延伸出一根長長的、由蛋白質組成的性線毛(Sex Pilus;F Pilus)。這根線毛會精準地結合在缺乏該質體的受體細菌(Recipient, F-)表面受體上。 2. 拉近距離(Retraction):一旦性線毛成功與受體細菌連結,這根線毛就會開始去聚合化(Depolymerization),也就是從基部開始拆解、縮短。這個動作會像收線一樣,將兩隻細菌強行拉近,直到它們的細胞膜緊密接觸並融合,形成一個穩定的接合通道(Conjugation bridge)。 3. DNA的單股切割與轉移(Transfer):遺傳物質(通常是質體 DNA)並不是整條直接送過去。供體細菌體內的酵素會在質體的特定位點(oriT)切開一個缺口,將雙股 DNA 拆開成單股。這條單股 DNA 會像穿針引線一樣,通過接合通道被「推」進受體細菌中。 4. 複製與完工(Synthesis):當單股 DNA 進入受體細菌後,兩隻細菌會各自以這條單股為模板,同步合成出互補的另一股,重新將其變成雙股螺旋的DNA質體。最終,原本的受體細菌(F-)也獲得了該質體,還可以正式轉變為擁有製造線毛能力的供體細菌(F+)。這也是為什麼抗藥性基因(例如對抗生素的抗性)可以在細菌族群中以驚人速度傳播的核心原因。簡單來說,細菌(真細菌域)線毛本身主要負責「抓取」與「拉近」,真正讓 DNA 通過的,是兩者細胞壁/膜融合後建立的線毛通道傳遞質體。這個機制繞過了傳統的有性生殖,讓細菌即使不細胞分裂,也能直接「水平分享」彼此的生存DNA資訊。 由於微生物界(細菌和古菌)的水平基因轉移(即透過質體、噬菌體直接交換 DNA之各類基因轉換方式)已瘋狂到超乎想像的程度。很多演化微生物學家多利托利(W. Ford Doolittle)提出強烈質疑與反思(註11):他認為在經過了 40 億年古代細菌先祖無休無止的水平基因轉移的基因大洗牌之後,早期的演化訊號可能早就演變成了一種「統計上無意義的隨機噪音(筆者註:指被水平基因移轉干擾到不具回溯之規律性)」。他也懷疑科學家現在用『超級電腦』推算出來『數位虛擬世界』中的 LUCA 基因集,有沒有可能根本不是歷史上真正存在過的那隻太古的祖先細胞,而只是電腦演算在面對無數次基因水平交換後,所產生的一種假設存在的「統計學幻覺(Statistical Artifact)」?』而把這些基因塞進同一個單一祖先細胞(單系群起源LUCA)」這樣的的想法可能是錯的。 由以上所述就發現:就連演化學者本身也有人對『親緣關係樹』的單一祖先LUCA感覺問題重重,信心動搖呢! 前提2:化學進化論的假定之中LUCA的分子鐘(Molecular Clock)的突變速率在40億年間必須具有跨時間的恆定性或可預測性:2024 至 2026 年的前沿研究(如前述 Moody 等人的 Nature 論文;註10)將 LUCA 的年代精準推算在 42 億年前。這種「親緣關係重建(Phylogenetic Reconstruction)」高度依賴進化模型的分子鐘假定:基因的突變速率可以透過某種統計模型(如鬆弛分子鐘模型)穩定向後回推。至終Moody 的權威自然雜誌論文承認 LUCA 在 42 億年前就擁有了 2,600 個基因和完整的 CRISPR 免疫馬達。這讓許多進化論學者與智慧設計論學者炸鍋了! 智慧設計論者曾提出LUCA分子鐘的前題提出質疑與指出盲點:在化學進化論的假定之中,42 億年前的冥古宙與地球環境與現在完全不同。化學進化論的假定之中當時沒有臭氧層,地表與海洋充斥著極高劑量的宇宙輻射與紫外線;且若依化學進化論者的預測早期的 DNA 複製酶(聚合酶)必定是非常原始,因子分子鐘的錯誤率必定極高。在化學進化論者的原始環境這種「高輻射、高突變率、缺乏精密修復機制(還沒演化出來)」的遠古時代,基因突變的速度可能比現代快上數百倍甚至數萬倍。如果當時的分子鐘走得極快,那麼科學家假設突變率恆定所推算出來的「42 億年」時間軸,就會產生巨大的誤差,LUCA 的存在年代和複雜度也會隨之瓦解。根據慕迪團隊分析LUCA的權威科學數據(註10),主流科學界普遍推測,生命的化學起源(從無機分子到第一組自我複製的 RNA 原始體)應該發生在43億至44億年前。這意味著,在地球形成後短短1至2億年內,生命就已經「開工演化」了。這不該走快的LUCA演化分子鐘卻突然異常走的極快,在1~2億年中竟然突然演化出2,600個相互合作精細分工的基因團隊,真的極其不可思議!這代表甚麼呢?凱西.盧斯金的專文指出(註16): 『進化論科學家也曾坦承:「在:LUCA一億年甚至更短的(分子鐘)時間內發 生如此巨大的進化,實在令人難以置信。」的確,進化非常劇烈。 諸如此類的轉變,包括非常迅速的轉變,是人類創造科技的標誌性特徵。就 早期地球而言,這聽起來像是某種創造性力量的行為,這種力量甚至在第一 個細胞出現之前就已經存在。換句話說,這聽起來像是智慧設計。 換言之,生命的高度秩序性已經被進化論學者自己的研究團隊刻劃出來了!異常快速的分子鐘似乎說出:若不藉著智慧設計者的力量,真細菌域與古菌域的諸多生命體是不太可能自己形成的! 智慧設計論根本沒有這許多進化難題要面對 但是如果三個域的所有物種都是被『智慧設計』的(如聖經創世紀第一章所述:萬物是各從其類而被創造),上述的許多廣進化理論的矛盾會自然消除。因為各自被設計的,各自有自己的起源,就可以各自擁有彼此相似與不相似的部分。在智慧設計的架構下三個域生物彼此之間的壁壘與鴻溝當然是理所當然會存在,也不需刻意尋找三個域之間彼此的演化上的關聯性。前寒武紀缺乏約30億年之久的大片進化過程中的化石空缺自然就符合聖經『各從其類』的觀點。化石群生物為何缺乏轉變的中間型的問題變為理所當然。接受智慧設計論不用硬著頭皮去解釋共同祖先A如何演化成B與C。也就是說,A、B與C物種均獨立源自同一位設計者。進化論的最大問題可能就是原先進化論者設定的『A演化成了B與C』的『廣進化框架』。就如凱特.拉希金(Casey Luskin)博士曾指出進化理論解釋的矛盾或許是肇因於廣進化本身(由簡趨繁的概念)的假設(如共同祖先A廣進化成B和C)可能就是錯的。智慧設計論的理論架構與微進化概念也可以並存,因此其合理性比廣進化理論架構的矛盾百出更是有過之而無不及!
1. 達爾文,物種源起,第十五章 複述和結論,電子書第319頁,谷月社電子書出版 2. 維基中文關鍵字(含2-6圖片來源):最後共同祖先 3. Madeline C Weiss 1, Filipa L Sousa 1, Natalia Mrnjavac 1, Sinje Neukirchen 1, Mayo Roettger 1, Shijulal Nelson-Sathi 1, William F Martin 1Nat Microbiol. Nat Microbiol. 2016 Jul 25;1(9):16116. doi: 10.1038/nmicrobiol.2016.116. The physiology and habitat of the last universal common ancestor. 數位物件識別碼:doi: 10.1038/nmicrobiol.2016.116. 4. Edmund R. R. Moody, Sandra Álvarez-Carretero, Tara Mahendrarajah, NIOZ Royal Netherlands Institute for Sea Research, James Clark, University of Bath, The nature of the last universal common ancestor and its impact on the early Earth system., July 2024, Nature Ecology & Evolution 8(9):1654-1666 數位物件識別碼:DOI:10.1038/s41559-024-02461-1 5. Caforio, A., et al. (2018). "Converting Escherichia coli into an archaebacterium with a hybrid heterochiral membrane." PNAS. 6. Casey Luskin, Is There Discontinuity in Biology — And How Would We Know? (2021) Science and Culture Today網路專文 7. In situ growth of the novel SM1 euryarchaeon from a string-of-pearls-like microbial community in its cold biotope, its physical separation and insights into its structure and physiology. Arch Microbiol. 2003 Sep;180(3):211-7. DOI: 10.1007/s00203-003-0580-1 8. , et al. 2015, S-layers at second glance? Altiarchaeal grappling hooks (hami) resemble archaeal S-layer proteins in structure and sequence. ORIGINAL RESEARCH article Front. Microbiol., 09 June 2015, Sec. Physiology and Metabolism of Microorganisms. Volume 6 – 2015 Doi: 10.3389/fmicb.2015.00543 9. Gauger, A. K. (2015). The Biologic Institute Reports: Lipid Membrane Chirality and the Limits of Enzyme Evolution. 10. Casey Luskin.(2024)“That Is a Lot of Evolution”: Study Finds LUCA Required 2,600 Genes., Science & Culture Today網路專文 11. W. Ford Doolittle, Phylogenetic classification and the universal tree. Science. 25 June 1999 Vol.284 (5423) p. 2124-2129.
|
|
| ( 知識學習|科學百科 ) |