|
 🌳進化論破綻大公開(一)出書❗ 🌳進化論破綻大公開(一)出書❗
沈瑞良老師自費出版的新書『進化論破綻大公開(一)』終於在2025/8/01正式出版了❗ 🌳在這本書封面寫著: 進化論是真理?或只是長久以來的社會和學術共識?地球上的生命可以自然產生嗎? ◎我們真是由「猿猴」與「智人」的共同祖先演化而來的嗎? ◎生物課本的進化論部分內容是真的嗎?抑或只是被定於一尊的假設? 這本書的封面說: ◎這不僅是一本科學論述的書籍,更蘊含著深刻的生命教育意涵。 生物進化的理論應該受到嚴格的科學檢驗, 本書不僅對進化論提出具體的科學質疑,更是一場對科學的求真探索! 生命是碰巧產生的,還是「智慧設計」的? 本書呈現「進化論破綻」的相關科學期刊論文重點,以及被忽略的科學證據。 另外,也將「智慧設計論」論證與推理以客觀角度呈現出來,讓讀者做同步的思辨與比較。 作者透過豐富的科學文獻與研究成果,試圖讓讀者重新審視種種被忽略或未被完整呈現的科學觀點,也鼓勵學術界回歸科學探索的初衷—— 不帶預設立場地追尋真相。 🌳作者介紹:沈瑞良老師,1963年出生,從高一開始就對「地球上的生命如何起源?」此一議題的追尋懷有極大的熱誠。 沈老師畢業於國立師大附中;中山醫學大學營養學系學士;中國文化大學生活應用研究所食品營養組碩士。曾任中華穀類食品工業研究所的副研究員2年,奎山中學生物教師兼任教學組長11年,之後並在台北市各高中擔任生物兼課教師長達13年之久至現今。曾任交響樂團小提琴手,亦為資深小提琴老師。 沈老師對與生命的起源有關的「進化論」與「智慧設計論」(舊稱創造論)一直有長久與深入的研究。繼1997年出版《科學否定進化⸺揭開進化論的疑點與破綻》(淑馨出版社,已絕版)之後,他多年來的研究結晶於本書再度有新的呈現。 🌳這本書還獲得者幾位醫界人士的重磅推薦: 1.本書試圖在這場科學與哲學的對話中,提供另一種值得深入思考的觀點。(林志慶/台北榮總腎臟科主任) 2.本書以詳盡查考的方法與正反並列的比對模式,給予讀者不一樣的思維邏輯。(董述平/澎湖醫院急診科醫師) 3.作者為我師大附中同學,一直以來認真研究生命的起源並探索人類來源,頗有心得,推薦之。(林誌文/大中牙醫診所醫師) 4.作者多年的執著,對「進化論」深入淺出地探索,值得理性品味,值得思源追溯。(林淳宏/腸胃科醫師) 5.演化論是人類智慧推衍出的理論,而學者們卻必須經常推翻、修改其中物種形成的一些重要理論,觀看本書可避免坐井窺天的狹隘。(張有德/高中生物教師) 🌳我出書的心路歷程: 出這本書其實跟我師大附中高一時的遭遇有關。記得我高一上生物課的時候,教材上到進化論的部份。突然老師就指著聖經的創世紀嘲諷了一番,說到:聖經創世紀泥巴人亞當,被吹一口氣就變成了一個人,如同女媧補天一般可笑。現今進化論才是科學的主流。老師的這番話使我頓時陷入困惑。到底是聖經的創造觀點比較正確,還是進化論猿猴祖先比較正確呢❓我頓時發現進化論其實牽動到這個宇宙與生命的起源,這會影響到我的人生觀,難道我不該搞清楚嗎❓但是我僅僅是一個懵懂的高一學生,實在很難自己從那個困局中出來。甚至到最後我也試著跟假設存在(也可能不存在)的宇宙主宰說話。我說:上帝啊!你到底是存在還是不存在,若是你存在,使我知道你的真實與進化論的錯誤;若你不存在,我現在只是在跟空氣說話,那反正我也不會有損失。 也許是這個禱告產生了功效,我接著就在逛書店的同時遇到了第一本智慧設計論學者的書,是旅美學者何天擇所寫的。他是一個留美的藥學博士,對進化論與智慧設計論的探索有連續16篇小論文,主要是站在智慧設計論的角度去批判達爾文進化論的『跨種演化』、『萬物同源與共同祖先』這部份的論點。我才知道進化論這部份只是一個假說而已。而且有許多科學證據並不站在進化論這邊,而且甚至是反證,最重要的,這些證據是課本上、報章雜誌上找不到,甚至是被消音的。 從何博士的論文引用證據都列有參考資料,我又抽絲剝繭找到了幾本美國幾位支持智慧設計論科學家的著作(他們都是博士級的社會精英): 杜安•季西的《化石否定進化》 亨利•莫瑞士的《科學創造論》(標題很吸引我,上帝的創造是聖經所主張的,卻仍舊能符合科學的證據) 伽利•帕克的《創造論---神話抑科學❓》 約拿單《進化論的聖像~~~課本教的錯在哪裏?》 Ratcliff的《老周的身體》[讀者文摘出版](這本書使我知到人體的精密、複雜、精巧,簡直簡直到了難以想像的程度,甚至一個人人體的血管長度就可以繞地球2圈半) 然而只讀這些書還其實不夠具有反駁進化論的能力。在一個很奇妙的機會下,我當了國中的生物老師,也在1993年完成了我的第1版的書:科學否定進化(淑馨出版社)。但之後我一直忙於工作就,停止探索進化論了,直至1999年左右我的一個非常熱心的讀者,或許他是上帝所派來的使者,他遠道而來我家敲門來找我,告訴了我美國生物化學麥克•貝西教授的這本巨著:達爾文的黑盒子(The Darwin's black box),而且把這本書送給我。我真的很謝謝他,因他喚醒了我研討進化論與智慧設計論的初衷,尤其是這本書中提到人體的血液凝固就需要同時啟動13個凝血機制,三個路徑都要成功不受到阻礙,血液才會凝固,而這些都是需要設計複雜的分子機器,而無法藉著漸近隨機的突變產生。 在2009年起我進一步當了高中生物老師,之後高一至高三選修生物的課程我也全部都教過。我也在網際網路讀到了更多智慧設計論者與進化論者的論文期刊資料,使我對進化論在廣進化這部份的破綻有更全面的瞭解,同時也發現了更多新的進化論相反的證據與論述。我開始重拾初心,有空的時候就把這些進化理論較明顯的『破口』加以整理,陸續在我的部落格『人是猴子演化而來的嗎❓』發表。 而這第2套書已出版的第1本----進化論破綻大公開(一),分別從六個層面,選出13個子題深入探索,期望能以平衡論述的角度,呈現在一般書籍與報章雜誌上不容易讀到的----關於進化論反面的期刊論文證據與論述。預計未來也要出版第2本與第3本,目前正在規畫撰搞中。 🌳內容簡介:在這本書的封面問了三個核心問題: 1.我們真是由,『猿猴』與『智人』的共同祖先演化而來的嗎❓ 2.生物課本的進化論部分的內容是真的嗎❓ 3.地球上的生命可以自然產生嗎❓ 在全國新書資訊網中對這本書的評述如下: 『 作者提出一個不同於主流教材的視角,希望透過對進化論論述中可能存在的謬誤進行剖析,來呈現另一種科學觀點。本書的撰寫,不只是對進化論的質疑,更是一場對科學求真的探索,提醒讀者在接受科學教育時,應該保持批判性思考的精神。』 在進化論破綻大公開第一章---進化理論矛盾破綻,提出論述的主要亮點是: 1. 進化論目前『只是假說』,卻被放在課本上被講得像真的一樣。這是由於絕大部份生物課本的課文是相信進化論的學者寫的,因此他們都會以進化論早就是『已成的事實』的口吻去敘述進化理論。事實上除了微進化的部份之外,進化論(主要是《共同祖先物種跨類演化》的部份)只是一個『尚待證實』的假設而已。 2.通常假設提出之後,需要以實驗驗證。但是廣進化竟然是是一個『無法被實驗驗證與推翻』的假設。因此進化論《共同祖先跨類演化》既無法被驗證;也無法被推翻。這是由於進化論所主張動輒數十萬年的演化時程,遠遠超過一般實驗設計的極限。 3.第一章內容還提到一個重要的論點就是:長頸鹿極長的脖子以進化論去解釋其成因,同樣是矛盾百出,並不比用進廢退說解釋的拉馬克正確。長頸鹿生存在非洲大陸的疏林草原,這是一個沒有高大的樹木的生存環境,與達爾文的演化理論產生嚴重的矛盾。 ------------- 在進化論破綻大公開第二章---化石反證與年代矛盾,提出論述的主要亮點是:課本上所說『化石支持進化論』是很有問題的: 1.呈現『大段的空白,極度地稀少』在5.42億『寒武紀大爆發』之前的廣闊地層,約佔進化史6/7。而這段34億年接近完全空白無紀錄的地層與進化論的預測不合,也可視為進化論的反證。 2.各類生物普遍性的缺乏演化的中間型化石,古生物的跨種演化時常跳tone找不到共同祖先。如地層中5000萬年前的蝙蝠化石(印手板圖1),與現代的鼴鼠(印手板圖2),這兩者看起來真的差很多❗進化論者認為古代鼴鼠跳tone成了古代蝙蝠,但無論是化石與現存生物都完全找不到中間型化石或生物,但雖然是如此,進化論者還是要『霸王硬上弓』說蝙蝠與鼴鼠這兩者有演化關係。 3.怪誕化石突然出現情況不少,且與前後化石均不連貫,造成演化論解釋的矛盾,如寒武紀的甲胄魚,在地層中突然出現,又突然消失。 因此照智慧設計論所預測的模型,這些地層中的化石應該是洪水所引發突發性的『快速照像』的被土石流掩埋,而比較不像是數億年的化石記錄,因此才未顯現緩慢改變的演化過程, ----------- 參、化學進化窒礙難行 1.生命起源的研究從尤里、米勒、福克斯之後並無進一步的突破——有機分子隨意的連接並不能產生生命體的高度的秩序性。首先出現的生命分子是RNA還是蛋白質⁉️兩者作為生命最初的物質都呈現了極大的障礙無法突破。 2.碰巧生成的小分子在稀薄的水溶液中具有分解的半衰期,在紫外線照射的不穩定環境中極其容易再被分解。 3.生命產生原始生命的過程中,小分子若自行連接為大分子是趨向低亂度(低熵),會違反熱力學定律[熵必需趨向增加]。由於生命體的產生必須是小分子要自行趨向整齊,這是違反生物熱力學第2定律的。因此生命的起源絕不會始自化學進化論所主張的『稀薄海洋中的熱湯的有機小分子自行連接(趨向熵的降低)』。 肆、人類演化問題多多 1.挖掘出來的『猿人化石』稀少、破碎又矛盾?——北京人、爪哇人有許多不為人知的祕辛(如爪哇人頭蓋骨與人類大腿骨埋藏地點相隔15公尺,但發現者杜布瓦卻硬是將此兩塊骨頭『判定』屬於同一個主人。) 2.科學期刊登載的300萬年前智人存在的證據,比直立猿人更早,但這些證據被進化論的媒體與教科書消音。 3.人類進化史約600萬年。但以人類人口的自然增加率,只要4千年便能由一對夫婦繁增到35億。猿猴演化成人的600萬年來『人口爆炸』竟然只有近代的這一次,這合乎常理嗎? ----------- 伍、分子生物演化障礙 1.黑猩猩與現代人『DNA相似度達98%』的說法,是進化論者以有利於進化假設的方法所測定出來。因此300個核苷酸小片段DNA的相似性與整個生物體的DNA的架構相似根本就是兩回事。 2.在2004年英國的自然期刊所登載出來黑猩猩21號與智人的22號染色體DNA的比較得到『差異很大』的結果,但這個結果被親進化論的媒體消音。 3.生物『染色體數』各不相同且差異極大,物種染色體數不能隨意增減,如何跨越染色體的障礙進行跨類演化?更何況有15600條染色體纖毛蟲這樣多染色體的精巧原生動物,形成進化理論無法解釋的情況! ---------- 陸、處處可見生物的智慧設計 1.視覺影像的呈像與神經傳導過程牽涉十個高度複雜的生物化學步驟,加上傳入大腦視區的完美路徑,呈現不可簡化的複雜性,達爾文曾經在物種起源這本書中坦承這是生物演化過程難以達成的。 2.美國生物化學教授主張:細胞內的分子機器處處可見,而『分子機器』是不可能進化漸進產生的。如簡單機械捕鼠夾共有五個零件組裝而成。而擁有1個零件與擁有4個零件結果是一樣的,都是不能捕鼠(如進化論的漸進的狀況)。因此細胞分子機器必須要開始就有全部的零件,並且必須將它們各就各位合式地組裝。這說出必然有一位智慧設計者的預先存在。 3.美國生物系教授迪恩•凱尼恩於1969年出版進化論著作之後,但卻在1974年在生命起源的研究過程中開始懷疑進化論,最後他在1976年轉而成為支持智慧設計論的倡導者,震驚美國科學界! ~~~~~~~~ 🍄本書60頁這段話可以視為這本書最後的結論,可以視為創造論與進化論之間的『平衡論述』: 🍄美國氣象學家多明尼克史坦森:『智慧設計論者並不認為原初上帝創造物種的是我們今天所看到的物種(智慧設計論修正了「物種固定性」,認同了微進化)……相反地,上帝創造了不同種類的植物和動物,並設計它們具有改變和適應不同環境的能力,有時甚至導致不同的物種(物種形成 )。然而,這種變化的程度被認為是有限的:水仙花可能變成其他種類的水仙花,或者馬可能變成其他種類的馬;但水仙花永遠不會變成蘋果樹(另一類植物),馬永遠不會變成長頸鹿(另一類動物)。』聖經中的創造萬物與達爾文所主張的微進化都是對的真理,但廣進化(共同祖先物種跨類演化)則缺乏證據,且與分子生物學證據及地層中的化石分佈情況抵觸。 🍄正如本書作者在序言中所描述的:進化論有真實與虛僞的兩部份——真實的部分與虛構的部分之區隔其實就是:『有限演化』(微進化;microevolution——真實的部分)與『大演化』(廣演化;Macroevolution——虛構或證據力不夠充分,卻假定為真,如海格的親緣關係樹)。 🍄若有意購買可參考以下網購售書點: 🌳博客來-進化論破綻大公開(一) 2025/8/14之前有79折優惠https://www.books.com.tw/products/0011026617?srsltid=AfmBOorSPv4tV06rgyaj9Vo6MEAjMksAMvqwzQBZYAR-nioFVia7IOsY ☘️誠品線上價85折 進化論破綻大公開 (一 )│https://www.eslite.com/product/10012084572682944917002?utm_source=app&utm_medium=share&utm_campaign=sku 🌳蝦皮:https://shopee.tw/product/84325322/43909361663/ 🌳PChome24H:https://24h.pchome.com.tw/books/prod/DJBP14-A900J38HR 🌳三民:https://www.sanmin.com.tw/product/index/014558168 🌳讀冊TAAZE:https://www.taaze.tw/products/11101068731.html 🌳MOMO網路書店:https://www.momoshop.com.tw/goods/GoodsDetail.jsp?i_code=14253614&Area=search&mdiv=403&oid=1_1&cid=index&kw=9786263646728 🌳墊腳石:已提供資訊給通路上架中,預計14-21個工作天 🌳金石堂:https://www.kingstone.com.tw/basic/2013620012917/?lid=search&actid=WISE&kw=9786263646728 |

|
文章數:33 |
 演化根源不明,代誌大條了?——我們是『古菌人』? 演化根源不明,代誌大條了?——我們是『古菌人』? |
| 知識學習|科學百科 2026/07/10 20:56:30 | ||||||||||||||||||||
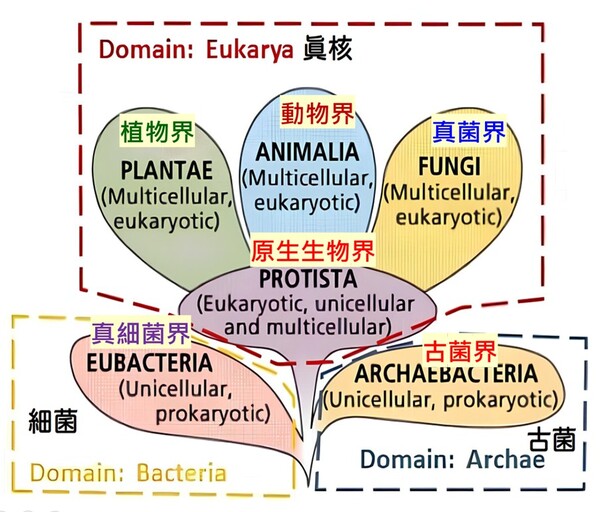
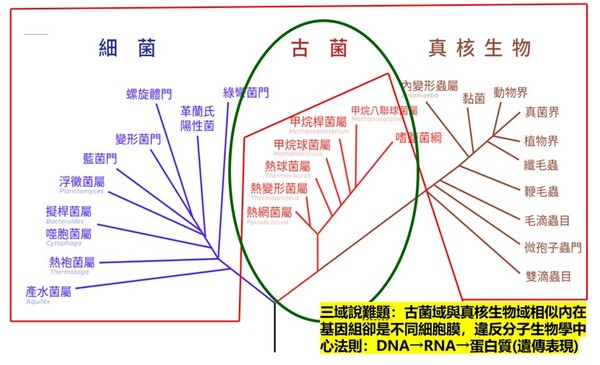
細菌可以演化成人嗎?進化論以親緣關係樹解釋萬物起源,但演化親緣關係樹的最基部的主幹竟是斷裂的?我們人類與地球上的生物最起初是從哪類生物演化而來的呢? 演化論學者藉『單一起源跨類演化』的親緣關係樹呈現各類別生物的演化關係,嘗試以DNA、RNA或蛋白質序列分析的相似性尋找分子生物學的演化證據,探尋原初地球上原始生命的共同祖先…… 但由於各演化學者探尋的角度不同,彼此的證據浮現了更多演化理論的矛盾!依照演化原理,地球上生命的基本類別到底要分為三個類群(兩小一大:真細菌域、古菌域與真核生物域【依此觀點我們算是真核生物域生物之一】),還是兩個類群(一小一大:真細菌域、古菌域【依此觀點我們算是古菌域生物之一,德州大學學者甚至稱呼人類為『古菌人』】呢?目前演化論學者缺乏共識相持不下,公說公有理,婆說婆有理,吵翻天了! 目前生物學界在生物物種的分類是以演化親緣關係由低等至高等的樹狀圖概念予以分類的。目前主流學術界所採用的生物分類系統,則是由演化論者作為「系統分類學家」(Systematists)的學術共識所主導與決定的。現代分類學的核心原則是演化親緣關係學(Phylogenetics),主要透過化石證據、分析DNA、RNA、蛋白質序列相似性或形態特徵,利用統計與計算模型建立各類生物跨種演化的「親緣關係樹」,來視覺化呈現物種的共同祖先及分支脈絡(註1)。其根本邏輯就是:「萬物都是演化而來的;因此分類必須反映各類生物演化歷史(單系群)」。這代表一個生物分類階層只要無法呈現廣進化上的演化親緣關係,在現代主流分類學中就會被視為「不合格」(例如:將鳥類與哺乳類皆歸納為『內溫動物(體溫較為恆定)分類群』的分類方式因主流的演化概念(羽毛恐龍化石)牴觸,就被判定為不符合演化原理的『多系群』。)至於合格還是不合格,則是要看這個分類群合不合乎演化學者的『主流共識』所裁定的演化途徑! 依演化觀點的「五界系統」(Five-Kingdom System)由美國學者懷塔克(Robert Whittaker)於 1969 年提出。該系統依據細胞構造(原核/真核)、細胞數目(單細胞/多細胞)以及營養方式(光合作用/分解吸收/攝食)將所有生物分為五大界,為目前生物學基礎教育中最常見的分類方式: 1.原核生物界(Monera):細胞中心為環狀DNA的原始擬核(Nucleoid), 無核膜,多為單細胞,代表生物為:細菌、藍綠菌(藍綠藻)。 2.原生生物界(Protista):細胞中心為真正的細胞核(Nucleus;有核膜(內含不同數目的線狀DNA),多為單細胞或簡單多細胞,代表生物為:草履蟲、變形蟲、眼蟲(以上為單細胞),以及大型藻類(多細胞,如海帶、紫菜)。 3.菌物界(Fungi):細胞中心為真正的細胞核,多為多細胞 (酵母菌除外),細胞壁含有幾丁質。無法行光合作用,靠分泌酵素分解至細胞外界產生有機物後再吸收 (異營吸收)。代表生物:蕈類(如香菇)、酵母菌、黴菌。 4.植物界 (Plantae):細胞中心為真正的細胞核,全數為多細胞的真核生物,細胞壁含纖維素。能利用葉綠體行光合作用自製養分 (自營)。代表生物:苔蘚植物、蕨類植物、裸子植物 (如松、杉、柏)、被子植物 (亦即開花植物)。 5.動物界 (Animalia):細胞中心為真正的細胞核,全數為多細胞的真核生物,細胞無細胞壁。具備運動能力,主要靠攝食來獲取養分(異營攝食)。代表生物:各類無脊椎動物(如刺絲胞動物的水母、節肢動物的昆蟲、蝦蟹)、脊椎動物(如脊椎動物的魚類、鳥類、哺乳類)等。而我們人類則歸屬於『動物界』的『脊索動物門』其中的脊椎動物(即以脊椎骨保護脊髓)。 直到1977年卡爾‧沃斯(Carl Woese)提出以『原核生物拆分』的觀點去把分類生物分類,後來也演變成「六界」的分類方式。依據核糖體RNA(rRNA)序列的差異,沃斯氏將懷塔克的原核生物界再增為兩個界而總共成為六界:可分為真細菌界、古細菌界、原生生物界、真菌界、植物界和動物界。沃斯氏在1990年再進一步將所有的生物再增加一個更高的階層:『域』,因此生物分類的八個階層由上而下分別是:域、界、門、綱、目、科、屬與種,域則共分為『三大域』,於1990年後獲得演化學者普遍接受(註2;圖2-4):
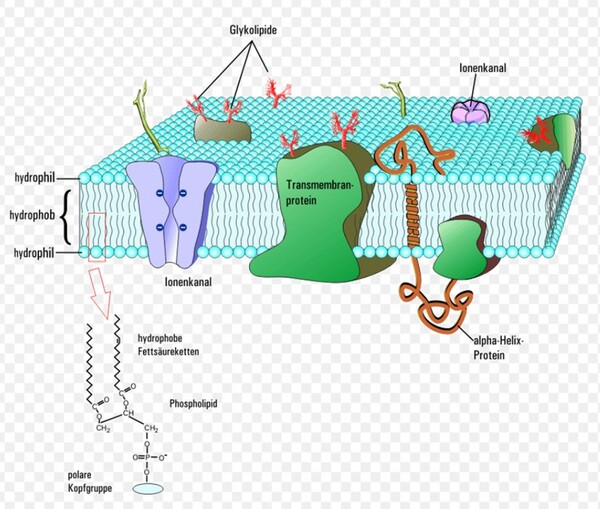
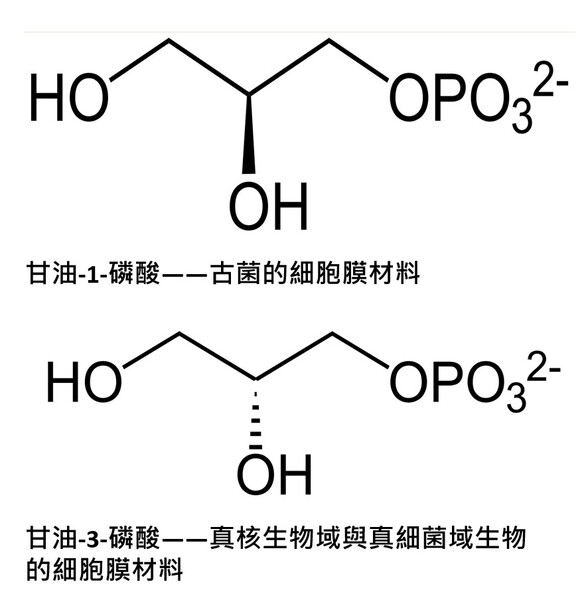
圖2-4 1990年後演化學者較普遍接受的三域六界系統分類圖,摘自咀嚼抹茶部落格 https://blog.gjlmotea.com/2082/%E7%94%9F%E7%89%A9%E7%89%A9%E7%A8%AE%E7%9A%84%E5%88%86%E9%A1%9E/ 2.古菌域 (Archaea):僅含一個古菌界,由真正的古菌組成,單細胞原核生物,但在基因結構上被演化論學者認為與真核生物更具親緣關係,常生存於極端環境,包含嗜熱溫泉菌、甲烷菌與嗜鹽菌。 3.真核生物域 (Eukarya):包含了所有擁有真正細胞核、膜狀胞器(如粒線體、葉綠體、內質網等)以及細胞骨架的單細胞或多細胞生物四個界(原生生物界、真菌界、植物界和動物界)。從微小的單細胞酵母菌,到巨大的藍鯨與神木,乃至人類自身,都屬於真核生物域。 三域說與二域說的棘手難題——細胞膜分子結構大不同的化學矛盾 真核生物域生物的四個界與真細菌域它們的細胞膜結構均是以『雙層甘油磷脂質』(甘油=丙三醇,具有三個碳原子)形成其細胞的重要基本架構(圖2-5左),而此種甘油磷脂質是以第3號碳原子連結磷酸基形成的2號碳『G-3-P手性』(甘油-3-磷酸手性,圖2-5右)。而古菌的細胞膜之構成材料的甘油2號碳則是『G-1-P手性』(甘油-1-磷酸手性,圖2-5右)。
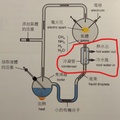
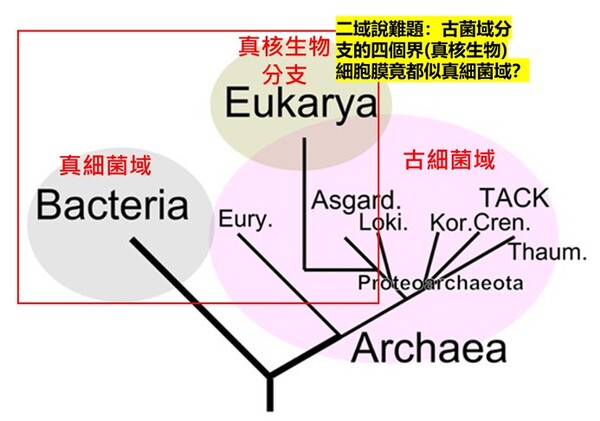
圖2-5(右) 在上半部(甘油-1-磷酸,Glycerol-1-phosphate,屬於古菌域細胞膜)的甘油磷酸分子結構圖中:2號碳(手性碳)的 OH 是黑色實心楔形(solid wedge)。在立體化學表示法中,黑色實心楔形代表鍵朝向觀察者,也就是「突出紙平面」。而在下半部(甘油-3-磷酸,Glycerol-3-phosphate,屬於真核生物域與真細菌域細胞膜)的甘油磷酸分子結構圖中:OH 是虛線楔形(hashed wedge)(凹入紙平面,遠離你)。另外兩條普通實線則表示位於紙平面內。圖片來源:維基百科關鍵字『細胞膜』,授權:CC BY-SA 3.0;『甘油-3-磷酸』授權:公有領域;英文關鍵字『Glycerol 1-phosphate』授權:公有領域。 1972年,美國加州大學的辛格和尼克森提出真核生物域細胞膜(真細菌域也相同)的流體鑲嵌模型(Fluid Mosaic Model)解釋細胞膜(或生物膜)的各種現象,是目前生物學界被最廣泛接受和認可的觀點。這種觀點主張:構成膜的蛋白質和雙層甘油磷脂類分子(甘油-3-磷酸結構)具有鑲嵌關係,而且膜的結構處於流動變化之中。隨後進行的研究有大量實驗結果支持流動鑲嵌模型的理論。在流體鑲嵌模型學說中,膜中的甘油磷脂質分子以雙層排列,構成了膜的基本結構。磷脂質分子為雙性分子,分為親水頭端和疏水尾端,雙層磷脂質分子之頭端(親水端)皆朝向細胞膜內側或外側的水相,因而使脂雙層分子之親水頭端面對細胞內與外,疏水尾端則兩兩相接埋於膜內。膜的另一種主要成分是蛋白質,蛋白質分子有的嵌插在脂雙層基本結構中,有的蛋白質貫穿細胞膜的兩側,有的則僅粘附於脂雙層的內外側表面上。這些蛋白質可能具有四種功能:1.運輸蛋白:跨膜的物質輸;2.酵素:協助細胞進行代謝;3.受體:與外界進行訊息溝通;4.辨識作用:辨別是否有「非自我細胞」入侵。 目前演化論學者對物種分類的看法上分成『三域說』與『二域說』兩大派系,這兩派其實都拿不出直接的化石證據,因此就致力發展DNA、RNA或蛋白質序列分析的相似性尋找分子生物學的演化證據,但是這樣的分子生物學分析在演化的不同學術機構的研究的不同解釋卻造成生物學界的『公說公有理,婆說婆有理』的『兩難』窘境。另有些時候不同演化途徑有時結果竟然分別演化出相似的生理結構(依演化原理不同演化分支應該生理結構也要不同)!這些窘境進化理論己乎都推給『趨同演化→因環境類似所以天擇演化出相似結構』這樣的說法: 1. 三域派 (Classic 3-Domain):由在1990年沃斯氏進一步提出,如以上所述:將演化親緣關係樹中所有生物物種都歸分於三大域的階層之下,即古菌域(Archaea)、細菌域(Bacteria)和真核域(Eukarya;即所有『有細胞核』的生物,包含動物界、植物界、真菌界與原生生物界)。進化論學者基於RNA資料的相似性,認為細菌、古細菌、真核生物三者的親緣關係如圖2-6所呈現的樹型關係:古細菌與真核生物擁有較晚近的共同祖先。 這一派遭受的困境是:無法解釋對於古菌域 (細胞膜構成為單層/雙層甘油醚、G-1-P手性、具有支鏈異戊二烯鏈)為什麼其核心基因組與真核生物類似,但真核生物域細胞膜模式(雙層磷脂質、G-3-P手性;圖2-5)卻又演化倒退與更原始的真細菌域(也是雙層磷脂質、G-3-P手性;圖2-5)相同。最近的全基因組定序技術科學家發現真核生物的「資訊系統」(DNA 複製、轉錄)像古菌,而「操作系統」(代謝、脂質)卻像細菌。這使得三域假說遭遇無法確定演化分支的極大挑戰(註3)。因為根據華生與克里克的『分子生物學』基因表現的法則:DNA(基因組)→RNA(抄寫密碼)→蛋白質(決定遺傳表現),真核生物域與古菌域既有『相似的基因組』,為何細胞膜的遺傳表現會變得極端不同,且甚至是完全不同原理、不同性質的兩種細胞膜系統呢? 如果分成三個域在親緣關係樹的演化會產生演化論者最不喜歡的細胞膜為雙層磷脂質的多系群(Polyphyly):又稱複系群:亦即依某演化分類群包含的生物成員跨越兩個或多個主流演化分支,但這樣的分類群關係卻與三域說的主流演化分支(由三域說演化學者所討論的親緣關係學主流共識形成)牴觸。不同演化枝幹怎麼最後會演化出相同的結構呢? 多系群的歸類為『反主流』的演化概念!換句話說,就是『生物類群的共同特性』竟然與主流親緣關係樹牴觸!也就明明就是『不同分支(不同單系群)』的生物類群,依主流的演化原理應該是隨機突變與隨機環境選擇,演化出來的構造應該毫不相似,但彼此之間怎麼竟然還是彼此有極特殊的部分竟然是很類似的!這樣在進化論者上只好說是『趨同演化(convergence)』,亦即環境又變得很類似,所以生物又演化成很類似了。殊不知這樣的推論又可能把進化理論又帶入循環論證(都是你在講的)的風險之中。例如:硬骨魚與人類(後口動物演化分支)和烏賊(前口動物演化分支)依演化原理兩類生物理應截然不同,但若將牠們的眼球切開剖析,你會發現它們竟然都完美具備了相機型眼球(Camera-type eye)的構造:
在嚴酷的海洋生存競爭中,要看清獵物和掠食者,這種「相機型」的幾何光學構造是最完美的物理學解法。於是,兩個完全毫不相干,分屬不同演化分支的物種,在相同的環境壓力下,竟然『無巧不成書』地各自摸索出了『同一套設計圖』!更何況廣進化親緣關係樹各類生物這樣例子還不少呢!然而若因此將烏賊的頭足綱與脊椎動物歸併為相機型眼球生物竟然要被視為多系群,違反主流演化共識。因此有學者指出:這樣看來,錯誤的可能是進化論本身那一套生物源自單一起源共同祖先,由簡趨繁的廣進化的假設! 約翰尼斯堡大學地質學博士凱特.拉希金(Casey Luskin)曾指出進化理論解釋的此種風險可能是肇因於進化論本身(單一起源共同祖先、由簡趨繁的概念)的假設(A廣進化成B)就是錯的(註4): 『當生物學家無法建構親緣關係樹時,他們常常會暫時訴諸其他過程來解釋那些不符合親緣關係樹狀模式的資料。其中一種解釋是『趨同演化』。演化生物學家假設生物在不同的演化譜系中獨立地獲得相同的性狀,而不是從共同祖先遺傳而來。每當演化生物學家被迫訴諸趨同演化時,這都反映出主要假設(指演化論本身)的失效,以及無法將資料擬合到樹狀模式。文獻中有很多這樣的例子,但幾個就足以說明(演化論本身有)問題了。』
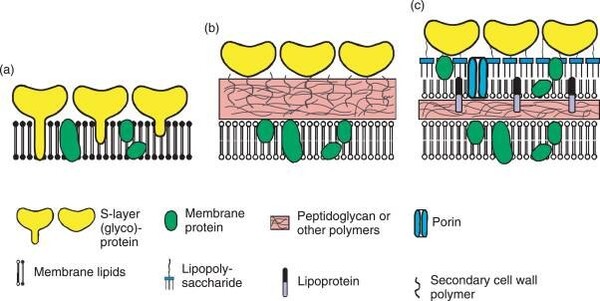
圖2-6 進化論三域派 (Classic 3-Domain;上圖) 二域派(2-Domain/Eocyte;下圖)與所遭遇細胞膜演化上的重大矛盾:紅色框內為細胞膜『雙層磷脂質,G-3-P,脂肪酸鏈』複系群分類生物;綠色框為細胞膜『單層/雙層甘油醚,G-1-P,具有異戊二烯支鏈』。 三域派的矛盾在於:為什麼真核生物的核心基因組【如同上游資訊系統】與古細菌類似,而細胞膜模式【如同下游操作系統】(雙層磷脂質、G-3-P手性、脂肪酸鏈)真核生物卻又趨同演化倒退到與真細菌(也是雙層磷脂質、G-3-P手性,脂肪酸鏈)相同。並且缺乏中間型態的化石。 二域派/融合派 (2-Domain/Eocyte): 認為真核生物是古細菌(阿斯嘉古細菌)的後代。但困境是無法解釋為什麼明明是不同的演化分支,細胞膜會在古菌(單層甘油醚、G-1-P手性、具有異戊二烯支鏈)進化為真核生物域的中途,細胞膜「砍掉重練」又被換回真細菌型(雙層磷脂質、G-3-P手性、脂肪酸鏈),並且缺乏中間型態的化石。 2. 二域派 (Classic 2-Domain): 二域系統是一種將演化親緣關係樹中的所有生物體分為兩個域的生物學分類:古菌域(包括真核生物)和真細菌域。它源自於對古菌多樣性認識的不斷深入,並對被廣泛接受的三域系統提出了挑戰。在1980年代,詹姆斯‧A‧萊克提出了始細胞假說,但由於當時的證據不足,該假說在很大程度上被三域系統所取代。21世紀初,人們對古菌,特別是它們透過與細菌共生在真核生物起源中所扮演的角色有了更深入的了解,這促使始細胞假說重新興起。特別是2017 年,日本科學家發現了一個名為普羅米修斯古菌(Promethearchaeati;舊稱阿斯嘉德古菌界Asgard archaea)的大型古菌界,似乎有進一步的證據表明,普羅米修斯古菌具有類似真核生物的蛋白結構(如RNA聚合酶與組織蛋白的相似性;膜變形與內吞作用的ESCRT-III 蛋白質系統),因此認為真核生物四個界應全數隸屬於『古菌域的分支』(註2)。科學家在 Asgard 古菌的基因組中,找到了大量過去被認為是真核生物特有的真核特徵蛋白質(ESPs, Eukaryotic Signature Proteins):細胞骨架(Cytoskeleton): 它們擁有編碼肌動蛋白(Actin)的基因,這能讓細胞改變形狀、長出突起。古菌的遺傳核心機制(DNA 複製、RNA 轉錄與蛋白質轉譯的分子機器),其與真核生物的相似度遠高於細菌。也使用一種類似組蛋白(Histones)的結構來纏繞包裝 DNA。依此觀點,在2023年甚至德州大學的論文戲稱我們人類乃是『阿思嘉德古菌人(Asgardians)』(註5)。 雖然基於細胞結構內部蛋白質的相似性,二域系統認為我們這些真核生物是古菌域的演化分支(即阿斯嘉德古菌是真核生物祖先的假說),但其演化上的困境是:無法解釋為什麼明明是不同的演化分支,為何細胞膜會在古菌(單層/雙層甘油醚、G-1-P手性、具有支鏈異戊二烯鏈)進化為真核生物分支的中途,整個真核生物的細胞膜被「砍掉重練」換成細菌型細胞膜(雙層磷脂質、G-3-P手性、脂肪酸鏈),並且缺乏中間型態的化石?細胞膜可是生物體非常重要的生命結構,其上還有數以千計的膜蛋白作為細胞的門戶,細胞膜又負責古菌與真細菌的能量代謝(筆者註:古菌與真細菌沒有粒線體)。如果親緣關係樹上,真核生物域只是從古菌域演化出來的一個分支,明明有相似的遺傳核心機制,但為何其細胞膜的主結構化學成分(雙層磷脂質、G-3-P手性、脂肪酸鏈)卻反而與『另一個域』(真細菌域)極相似,而不是與古菌域相似呢 (如圖2-6) ?換言之,二域說的演化途徑仍是擺脫不了為何古菌域的『真核生物分支』其細胞膜最後卻演化為與另一真細菌域相同的『雙層磷脂質細胞膜』的嚴重矛盾(與二域說的主流演化分支牴觸)! 我們目前看到的 Asgard古菌(如 MK-D1),全身上下都還是百分之百的古菌醚鍵膜。它們體內雖然有真核生物的蛋白質(ESPs),但膜的生化結構卻沒有半點要「變成真核生物」的跡象。 這也是為什麼許多抱持批判思維的學者認為,單憑「基因相似度」就直接斷言人類是古菌人,有些過度簡化了細胞生化演化的複雜度。要跨越這道『脂質大峽谷』,需要的不是隨機突變,而是兩套完整且不相容的生化微工廠是否進行了完美的交接。但是目前這樣的交接在化石與現存生物都還沒看到一點蛛絲馬跡! 真細菌與古菌細胞膜與細胞壁結構矛盾『水火不容』顯示『生物並非單一起源』 儘管二域說在進化理論發展上逐漸佔上風,但進化論學者在科學論文中仍坦承二域說這其中細胞膜與細胞壁結構在演化過程如何由單一原始生物起源逐漸演變過程幾個大的嚴重難題: 1. 兩者生物膜物理特性極不相容,不可能混合存在:古菌域細胞膜 (單層/雙層甘油醚為基本構成)與和真細菌域細胞膜 (雙層磷脂質)兩種『磷酸甘油骨架』手性(chiraty)截然不同,很難混合成穩定的生物膜而變成混雜的四不像,因特性不明確,最後在功能上一事無成。進化論者卡弗里歐坦承(註6): 『細菌和真核生物的膜脂由脂肪酸透過酯鍵與甘油-3-磷酸(G3P)連接而成,而古菌的膜脂則具有異戊二烯基烷基支鏈,這些烷基鏈透過醚鍵與甘油-1-磷酸(G1P)連接,導致甘油磷酸骨架的立體化學結構相反。這種「脂質矛盾(Lipid Divide)」引發了人們對微生物生命演化的疑問,因為真核生物被認為起源於古菌,這需要膜組成發生根本性的改變。』 進化論者維克托.索喬(Victor Sojo) 在2014年的研究也指出(註7): 『古菌域和真細菌域是親緣關係樹最底層的分支。這兩個類群在形態上相似,並共享一些基本的生物化學特徵,包括遺傳密碼,但它們之間的差異卻十分顯著,堪稱生物學中尚未解決的重大難題之一。這兩個類群的細胞膜和細胞壁組成截然不同,而DNA複製機制似乎也毫不相干。……古菌域和真細菌域在碳和能量代謝、以及膜脂立體化學方面的巨大差異,反映了不同族群中主動運輸作用的兩種獨立起源。……它們在細胞膜和細胞壁、糖酵解、離子主動運輸甚至DNA複製等基本特徵上存在差異。 兩者細胞膜脂質的差異可能是解決生物學中這一重大未解難題的關鍵。古菌的磷脂側鏈通常是異戊二烯類化合物,而細菌的磷脂側鏈通常是脂肪酸。雖然這可能反映了適應性演化,但古菌和細菌在甘油磷酸頭基的立體化學結構上也存在差異。古菌脂質具有sn-甘油-1-磷酸(G1P)頭部官能基,而細菌則使用鏡像結構sn-甘油-3-磷酸(G3P)。目前尚無令人信服的選擇性解釋來說明這些相反的立體化學結構。而相關的酶,古菌中的甘油-1-磷酸脫氫酶(G1PDH)和細菌中的甘油-3-磷酸脫氫酶(G3PDH),在系統發育上也沒有相似性,表明它們是獨立起源的。』 2. 細胞膜如何在演化的過程結構大換血? 宿主古菌必須在進化為真核生物的某段時間之內,把自身的古菌細胞膜(單層/雙層甘油醚、G-1-P手性、具有支鏈異戊二烯鏈)完全替換成與原先完全不同的細菌細胞膜(雙層磷脂質、G-3-P手性、脂肪酸鏈),且在這個演替過程中細胞不能崩潰而死亡。但這聽起就像是神話故事。因為在更換細胞膜的演化過程中,稍有不慎,細胞就會因為滲透壓失衡或膜蛋白失效而直接解體。這還沒有考慮真細菌與古菌細胞膜上還有各種特殊的膜蛋白,兩類生物各自不同的酵素系統如何相容呢!進化論學者有試圖提出可能的演化過程的各種猜測,但是仍是屬於各說各話尚無定論的階段!兩種細胞膜結構矛盾,水火不容,誰演化成誰都不對,這樣豈不顯示『生物應非單一起源』呢? 3. 細胞壁為何彼此迴異? 古菌域與真細菌域兩種不只細胞膜非常不同,細胞壁相較兩者也是結構截然不同(圖2-9;註8)。舉例而言,古菌域的細胞壁完全不含真細菌域細胞壁的肽聚糖結構,而是S層(晶格狀蛋白質)或假肽聚糖外覆結構的細胞壁為主,而真細菌分革蘭氏陽性菌與陰性菌。陽性菌具有厚的肽聚醣外層細胞壁,有的陽性菌也會在外覆S層結構,革蘭氏陰性菌具有薄的肽聚醣外層與甘油磷脂雙層的外膜的複合細胞壁。這些截然不同的特徵都代表這兩種域的物種其實應該沒有演化關係。再者,革蘭氏陽性菌與革蘭氏陰性菌的細胞壁也是彼此大不同(圖2-9)!
|
||||||||||||||||||||
| 最新創作 |
|
||||
|
||||
|
||||
|
||||
|
||||
| 最新影像 | 2 本 196 張 |
|
|
|
|||||||||