字體:小 中 大
字體:小 中 大 |
|
|
|
| 2024/02/08 01:12:23瀏覽6394|回應5|推薦0 | |
|
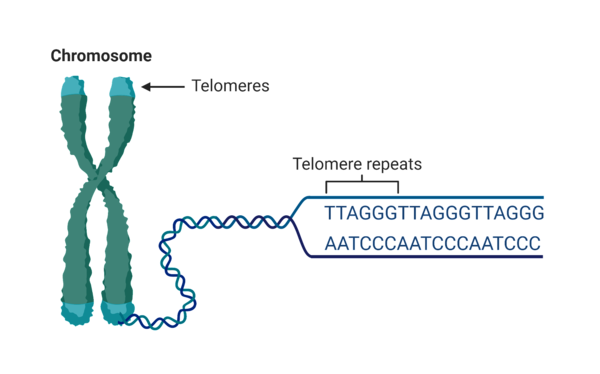
黑猩猩一直被認為是與人類親緣關係最近的生物,而且在進化論者的認定中,人與黑猩猩是有共同祖先的。他們認為現代人高度進化,而黑猩猩則沒有甚麼大幅度的進化,所以兩者在600多萬年前各自進化分道揚鑣。按照這個假定,這個共同的祖先(古人猿)就應該是和黑猩猩(包括猩猩、大猩猩和紅毛猩猩)極度相似,有24對染色體。而一般不同生物物種之間具有種別性,很重要的原因是:不同物種具有不同的染色體數,如獅、虎、家貓均有38條、野生小麥有14條、浣熊為38條、捲尾猴則為54條、而菲律賓眼鏡猴(Tarsiers)的染色體數更多達80條(註1)。染色體數不同因而就成了各類生物在演化上難以跨越的遺傳障礙。 然而若人類真是源自於古猿類祖先(先假定其為48條染色體),在演化過程要如何減少細胞核內的染色體,發生染色體突變而進化為現代人(46條染色體)便成為演化論學者中的一個極難解的議題。於是便有進化論者提出了黑猩猩兩條較短的染色體(第12與第13號,分別被命名為2B與2A段)因產生染色體突變的『羅勃遜易位症』融合為一條人類的2號染色體的說法(號碼越小的染色體為較長的染色體)。如右圖1是黑猩猩 2A 和 2B 染色體端到端融合形成人類 2 號染色體的假設模型。根據 Yunis 和 Prakash 兩位進化論學者發表的細胞遺傳學圖像按比例繪製染色體(註2)。請注意大小差異,若以上假說為真,基於人類 2 號染色體的已知大小,該差異約縮小了 10% 或 2400 萬個鹼基。而1991年一些相信進化論的研究人員在人類第 2 號染色體上發現了它們認為可能的融合事件的第一個所謂的DNA融合端(註3;Fusion site),然而這也就僅是一個僅含798個鹼基配對的『小的』、『混亂的』『似端粒樣』末端序列段,它們只是模糊地被進化論者類似於可能的融合。 至於原有黑猩猩的11號染色體也被論文以『融合必定成功』的觀點而稱作『2A染色體』(人類2號染色體A段) ,其中節區域就自動成為人類2號染色體的中節(右圖左上角);而12號染色體(被論文稱作2B段)其上異染色質的非編碼『中節片段DNA』也被認為自動退化消失而以演化觀點稱作『隱密性中節(Cryptic centromere)』(右圖右下角)。 就僅僅只是上述一篇期刊的推測,主流的進化論媒體就立刻加以大幅報導聲稱:科學家已找到黑猩猩的2條染色體融合為人類染色體的證據了!黑猩猩古人猿進化成為現代人是可能的!進化論者振奮不已,口耳相傳!然而,事實的真相果真是如此嗎? 一般染色體末端有很長的端粒區塊防止染色體彼此輕易融合 遺傳學者都知道真核生物染色體兩端的末端均有端粒(Telomere;註4)的結構。端粒是DNA重複序列的非編碼DNA(如不斷重複的TTAGGG) 的六鹼基重複序列,在染色體末端一遍又一遍地重複形成的區塊。其作用是保持染色體的完整性和控制細胞分裂週期。 由於DNA複製的機制,每次染色體複製後,延遲股上的染色體末端必無法被複製。因此,真核生物在染色體末端的端粒作為可被重複遺棄的片段。一旦端粒消耗殆盡,細胞將會立即啟動凋亡機制。因此,端粒被推測和細胞衰老有明顯的關係。人體的部分細胞,例如精原母細胞(精原細胞)、癌症細胞等,含有端粒酶,能在DNA末端接上新的端粒片段,其端粒不會隨著分裂次數增加而縮短,因此能無限複製。試問:黑猩猩的染色體既然有端粒的保護作用,怎麼會無故隨意融合在一起成為一條染色體呢?
黑猩猩染色體若丟失端粒產生融合,這與人類的羅勃遜易位症的典型現象並不符合 由生殖醫學權威醫師陳俊凱的部落格(註9)可以知道:人類的羅勃遜易位症較多發生於具有短臂(多半不含重要基因)的染色體,包含第13、14、15、21、22號染色體,這些染色體在短臂附近的著絲點融合時丟失短臂時不會造成重要的基因丟失。而黑猩猩染色體丟失端粒產生融合的第12、13號染色體並非是此種短臂染色體!因此黑猩猩染色體融合假說與人類實際的羅勃遜易位症病例典型(中節附近發生)並不符合。精通遺傳學的學者張維克在著作裡曾指出此一假設不符真實現況的重要關鍵: 『另外,〝高帽子〞(端粒)的作用,本來就是防止染色體互相之間發生融合的,恰好發生在“高帽子”上的融合,怎麼是可能的呢?在人類羅伯遜易位的染色體病例中,會發生兩個染色體的融合,染色體的斷裂處都是在中節的位置或中節附近的位置,端粒被丟失了,才容易發生融合。如果端粒沒有丟失,是不會發生融合的,因此(人類病例)沒有見到發生在端粒處(頭對頭)融合的病例。』(註5) 而大多數的人類羅勃遜易位症患者染色體是奇數45條(註10),子代高比例是不孕症(減數分裂精子或卵將少一條染色體,使精子或卵死亡)。試問:進化論者相信此種弱勢的遺傳疾病患者其子代要產生『進化的強勢物種』,這樣的邏輯是合理的嗎? 染色體融合位點僅798鹼基對,尺寸過小不合常理 然而如右圖上(註6),精通染色體結構的學者張維克提到一般染色體末端的端粒DNA序列應有500~2000個鹼基對。如果進化論者認為黑猩猩的兩條的12、13號染色體的端粒可以『頭對頭相接』,則相接後黏合的端粒區塊應該有1000~4000個鹼基對才對!然而進化論學者宣稱在人類2號染色體上找到的融合部位(fusion site)竟然只有798對鹼基對,這顯然並不符合兩個端粒相接後的正常大小。進化論者所主張的『驟變猿人』若存在,其丟失的端粒鹼基對區塊不應該不合理地無故消失才對。且年輕人與老年人精子的端粒標記序列並無不同,老年人的精子端粒並不會變少,受精卵形成的子代2號染色體既是融合的染色體,而其端粒的標記序列TTAGGG卻極少,這與『驟變人猿』染色體融合的進化理論不是嚴重矛盾嗎?遺傳學者克萊姆森大學教授湯姆金博士(Jeffrey P. Tomkins;註7)的網頁專文也指出此一嚴重問題: 『另一個問題是人類2號染色體(所謂的)融合位點很小,只有798個DNA字母長。一般染色體末端的端粒序列長 5,000 至 15,000 個鹼基。如果兩條染色體融合,您應該會看到 10,000 到 30,000 個鹼基長的融合端粒特徵,而不是798個。』 綜上所述,進化論者主張的黑猩猩染色體『頭對頭相接』的假定,其實是極度違反自然現象且極不合常理的。 798鹼基對融合區塊端粒標記序列零散、不連續又稀少 令人訝異的是:所謂的融合標記798鹼基中含有的端粒標記序列TTAGGG才只有52組【上圖(註6)鹼基對108305至108305的區塊】!端粒標記序列其實在所謂的2號染色體融合區域其實是零散、不連續又稀少的: 『與這些內部端粒標記序列相關的一個重要屬性是它們主要是單獨存在的。在融合位點兩側的 52 個完整 TTAGGG 標記序列中,僅發現 3 個串聯(筆者註:正常的端粒有約830至2500組連續大塊串聯)出現,其餘以獨立單體存在。在融合位點兩側的 154 個完整 CCCTAA 標記序列(筆者註:因DNA為雙股鹼基配對,AG與CG為配對關係,因此CCCTAA是TTAGGG其負鏈的部分的端粒標記序列)中,發現了 18 個串聯的標記序列,其餘序列為獨立存在。儘管標記序列和二聚體重複的密度在推定的融合區域附近有比較高,但它們在(筆者註:DNA序列的)閱讀框架中從一個 6個鹼基的端粒重複到下一個的位置卻是飄忽不定(且可能已不在框架內)。 由於端粒重複序列極其稀少,其大部分為單體狀態,並且普遍存在於假定的融合位點兩側,因此幾乎沒有數據表明它們可能曾經形成 10 至 15 kb 的完美串聯序列的6 個鹼基重複。進化論者所謂融合附近區域的2qfus 其序列明顯退化超出了顯示完整端粒曾經存在的程度。(註13)』 如以上所述,理應存在的大塊6鹼基端粒標粒序列在人類的2號染色體有『集體失蹤』的現象!試問:這與『驟變人猿』染色體融合的進化理論不是有嚴重矛盾嗎?試問:進化論者還要堅持所謂『頭對頭的端粒融合』使黑猩猩演化成現代人類嗎? 融合的染色體丟失1/3長度,竟能增加794個基因? 此外,根據張維克、宋國平所著——『依據DNA編碼的物種起源學』這本書中也提到:若黑猩猩染色體融合真的曾經發生,以融合長度而言,融合的人類2號染色體在融合過程等同丟失1/3的DNA長度,卻莫名其妙反而增加了794個基因,以基因總數而言,是DNA大幅減少長度,卻反而增加基因,這根本是極其不符合遺傳原理的:
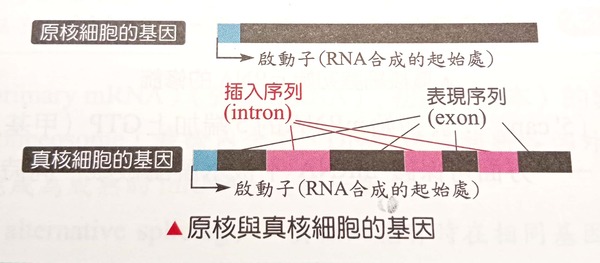
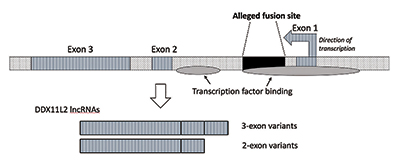
黑猩猩隨機形成的融合標記竟可成為現代人的基因開關——啟動子? 湯姆金教授根據他的研究更進一步指出對融合假說更具殺傷力的證據:2號染色體隨機產生的798鹼基對融合位點現在被證實是現代人DDX11L2 RNA解旋酶基因的開關——啟動子的一部分,屬於基因的『內含子』段落。這也太奇怪了!進化論者的融合假說認為因染色體突變的隨機接合的融合位點,竟位於現在稱為DDX11L2的 RNA 解旋酶基因內有其重要功能(註7) !所以『染色體不小心接在一起了,卻神奇地忽然形成了細胞重要功能零件』,這種歪打正著的邏輯是合理的嗎? 然而到底什麼是『啟動子』呢?首先要認識真核生物基因表現的方式: 真核生物大部分基因(含具有遺傳密碼的編碼DNA)的其中一股(DNA為雙股螺旋長鏈結構)經RNA聚合酶轉錄後會產生初始訊息RNA(mRNA),這是DNA用以製造蛋白質的初始RNA核苷酸序列,如此可將遺傳密碼抄寫至訊息RNA之上,攜出至細胞質用以主導其內蛋白質的製造。而轉錄必須從基因的『啟動子』區塊開始;也就是說,啟動子是RNA聚合酶的轉錄因子附著的起始位置。而轉錄因子可以影響RNA聚合酶的轉錄。而真核細胞的基因產生的訊息RNA其序列大多間斷不連續,由『表現序列(外顯子)』與『插入序列(內含子)』相間構成(如右圖註11)。表現序列為與製造蛋白質有關的序列,含有許多具有遺傳密碼的編碼區。插入序列為與製造蛋白質無關的序列,含有許多不具有遺傳密碼的非編碼區,且安插於各表現序列之間。
上述之初始RNA經過剪接修飾去除插入序列之後形成成熟的RNA,成熟的RNA進入細胞質中其密碼可被核糖體讀取,進而製造各種蛋白質,進行遺傳表現與產生結構與生理功能。 然而最奇怪的就是:所謂的黑猩猩兩條染色體融合位點既是染色體突變『瞎拚瞎湊的產物』,怎麼竟然會變成為一個DDX11L2 RNA解旋酶基因的重要開關——啟動子的一部分呢?正如湯姆金教授在他的專文中強烈主張:所謂的黑猩猩羅勃遜易位所產生的染色體頭對頭相接『融合位點』,絕不可能因此種災難性事件而成為開啟基因功能的『意外副產品』(註7): 『當我進行進一步研究時,我證實融合位點位於現在稱為DDX11L2的RNA 解旋酶基因內。植物和動物中的大多數基因都有其編碼片段,稱為外顯子,因此它們可以選擇性剪接。基於外顯子的添加或排除,基因可以產生多種產物。外顯子之間的介入區域稱為內含子,通常包含控制基因功能的各種訊號和開關。所謂的融合位點位於DDX11L2基因的第一個內含子內。 DNA分子是雙股的,有一條正鏈和一條負鏈。它的設計方式是為了最大限度地提高資訊密度,同時提高效率和功能。結果,相對的鏈上有不同方向的基因運行。事實證明,DDX11L2基因是在負鏈上編碼的。由於人類基因就像瑞士軍刀一樣,可以產生多種 RNA,因此DDX11L2基因會產生由兩個外顯子組成的RNA短變異體和由三個外顯子組成的RNA長變異體。 融合位點竟可以是基因啟動子嗎?這個DDX11L2基因可能有什麼作用呢?我的研究表明它在至少255種不同的細胞或組織類型中表達。它也與多種其他基因共表達(同時打開),並與細胞外基質中的細胞信號傳導和血細胞生成相關的過程相關(註12)。所謂的『融合序列』竟是位於與多種細胞過程的遺傳學相關的功能基因內,這強烈駁斥了它是頭對頭端粒融合的意外副產品的觀點。這說出基因不該是由災難性的染色體融合形成的! 更令人驚訝的是,融合位點本身具有功能性的,並且具有重要的工程目的。該位點實際上充當控制基因活性的開關。在這方面,大量生化數據表明,有12 種不同的蛋白質(稱為轉錄因子)調節該基因片段。其中之一正是 RNA 聚合酶 II,它是在轉錄過程中從 DNA 複製 RNA 分子的主要酵素。進一步支持這項發現的事實是,實際的轉錄過程是在所謂的融合位點區域內啟動的。』 綜上所述,『染色體不小心接在一起了,卻神奇地忽然形成了細胞重要功能零件』這件事本身就是極其不合邏輯的。正如我們家中浴室的蓮蓬頭,書房的門把都不該是以錯誤與隨機的方式碰巧製成的,你說是嗎? 染色體數目一樣,猿就可以演化成智人?進化論者把問題想得太簡單 再者,難道染色體數目一樣,猿類就可以變成智人?其實智人與猿類(包含黑猩猩)在數以萬計的『基因表達』上的極大不同,恐怕才是造成彼此生命表現顯著差異的重要原因。曾任美國獸醫部官員的珍.萊特納(Dr. Jean Lightner)博士也指出進化論者顯然把問題想得太簡單了!智人與猿類的細胞在基因的表達上有顯著的不同,其中許多的過程是藉著『編碼 microRNA (miRNA) 』對其細胞的調控,這不單純只是藉著染色體數演化成一樣都是46條這麼簡單而已!這就是為何猿類不會說話,而智人卻可以說出很清楚的語言(註14):
『人類在認知和語言能力方面與其他動物明顯不同。有時,黑猩猩使用工具或簡單手語的能力被吹捧為它們與我們關係密切的證據。事實上,黑猩猩在這些方面與許多其他哺乳動物和鳥類並沒有顯著不同(除了它們可以像我們一樣使用雙手)。黑猩猩缺乏人類語言的解剖結構。諷刺的是,有一些鳥類能夠很好地使用人類語言,至少對動物來說是如此。』
『儘管人類和猿染色體表面上相似,但在分子層面上存在重要差異。人類有許多蛋白質編碼基因是人類特有的,在黑猩猩中卻沒有發現。也許更顯著的是不編碼蛋白質的基因的差異。編碼 microRNA (miRNA) 的基因已被描述。 miRNA 分子不會被翻譯,而是直接控制基因表現。單一miRNA可以調節數十甚至數百個基因的表達。一項對大腦中表達的 miRNA 的研究發現,447 個新 miRNA 中有 51 個明顯是人類的,25 個只在黑猩猩中發現。(註15) 有如此多的基因被改變以便這些細胞是根據細胞類型以適當的物質濃度表達,使這些細胞可以有效地控制它們所調節的許多不同基因,這樣精細的機制並不是我們所期望的偶然過程能夠完成的。(註16)』 ………………………………………………………………………………………………………………………….. 羅勃遜易位症雖然能勉強解釋病態黑猩猩祖先48條染色體如何減少2條染色體形成46條,但是衍生出來的問體卻更多,也不符合新的遺傳學研究結果。試想:羅勃遜易位症的精子與羅勃遜易位症的卵子,丟失染色體臂與基因而成為受精卵,應產生病態古猿人,結果竟然沒大幅退化卻反而成為有高度進化優勢的現代人——此點極度不合邏輯!再者智人與猿類的細胞在基因的表達上藉『編碼 microRNA (miRNA) 』造成對其細胞的調控也有顯著的不同。生物在演化過程欲跨越染色體障礙,其本質上應該就是極其困難的。 更何況:
綜合以上所述:黑猩猩與人類既然有染色體數之差異及以上重大特徵之九大不同點,進化論者這樣勉強推測黑猩猩古猿人可進化成現代人類,其中的矛盾可真的是罄竹難書。 §參考文獻與附圖
11.施懿修編著:高中生物總複習(下)整理篇,華逵出版社,p.259~260 12. Tomkins, J. P. 2013. Alleged Human Chromosome 2 “Fusion Site” Encodes an Active DNA Binding Domain Inside a Complex and Highly Expressed Gene—Negating Fusion. Answers Research Journal. 6:367-375. 13. Jeffrey Tomkins and Jerry Bergman, 網頁專文The chromosome 2 fusion model of human., evolution—part 2: re-analysis of the genomic data, https://creation.com/chromosome-2-fusion-2
|
|
| ( 知識學習|科學百科 ) |